Sanne mekanismer til komplekse organismer
Oversatt herfra. {Kursiv og understreking er lagt til av undertegnede.}
Genreguleringsnettverk
Epigenetikk refererer til arvelige endringer i genuttrykk som skjer uten endring i DNA-sekvens. Disse endringene kan induseres spontant, induseres av miljøfaktorer eller som en konsekvens av spesifikke mutasjoner. Det er to primære og sammenkoblede epigenetiske mekanismer: DNA-metylering og kovalent modifisering av histoner. I tillegg har det blitt tydelig at ikke-(protein) kodende RNA også er intimt involvert i denne prosessen. De forskjellige mekanismene som kontrollerer epigenetiske endringer står ikke alene, og det er en tydelig sammenkobling og gjensidig avhengighet dem imellom:

Celletype og mønstre funnet i dyreriket genereres ved følgende signalveier:
- Hedgehog(Hh); - Vingeløst relatert; - Transformerende vekstfaktor-β (Tvf-β)
- Reseptor-TyrosinKinase (RTK); - Notch; - JAnus Kinase (JAK)/signalomformer
- AktivaTorer av Transkripsjon-proteinkinaser; - Kjernehormonveier
- Morfogenetiske proteiner; - Epidermale vekstfaktorreseptorer
- Fibroblastvekstfaktorer; - DNA-metylering
- Histonmodifisering og inkorporering av histonvarianter
- Kromatin-ombygging i eukaryote celler; - Ikke-kodende RNA-mediert epigenetisk regulering
24 Epigenetiske koder:
Den genomiske regulerings kode; Spleisekodene; Metabolsk kode
Signal-overføringskodene; Signal-integrasjonskodene; Histonkoden
Tubulin-koden; Sukkerkoden; Den glykomiske koden
Den ikke-ribosomale koden; Kalsiumkoden; RNA-koden
En domene-substratspesifisitetskode for NonRibosomal PeptidSyntetaser (NRPS)
DNA-metyleringskoden; Koaktivator/korepressor/epigenetisk kode
Transkripsjonsfaktorkoden; Modifiseringskoden etter translasjon for transkripsjonsfaktorer
HOX-koden; Den synaptiske adhesive kode; Apoptosekoden; Ubiquitin-koden;
Den bioelektriske koden; Transkripsjonell cis-reguleringskode;
Den nevronale pigg-koden
Gener involvert i celle-celle kommunikasjon og transkripsjonskontroll er spesielt viktige for dyrs utvikling
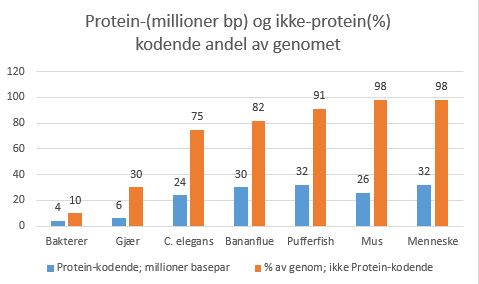 Hva er gener dyrene deler med hverandre, men ikke med andre riker? Disse kan forventes å omfatte gener som er spesielt nødvendige for dyrs utvikling, men ikke er nødvendige for unicellulær eksistens. Sammenligning av dyregener med genomet av spirende gjær - en encellet eukaryot - antyder at tre klasser av gener er spesielt viktige for flercellede organismer.
Hva er gener dyrene deler med hverandre, men ikke med andre riker? Disse kan forventes å omfatte gener som er spesielt nødvendige for dyrs utvikling, men ikke er nødvendige for unicellulær eksistens. Sammenligning av dyregener med genomet av spirende gjær - en encellet eukaryot - antyder at tre klasser av gener er spesielt viktige for flercellede organismer.
i. Den første klassen inkluderer gener som koder for proteiner som brukes til celle-celle-adhesjon og celle-signalering; hundrevis av menneskelige gener koder for signal-proteiner, celleoverflate-reseptorer, celleadhesjonsproteiner eller ionekanaler som enten ikke er til stede i gjær eller til stede i mye mindre antall.
ii. Den andre klassen inkluderer gener som koder for proteiner som regulerer transkripsjon og kromatin-struktur: mer enn 1000 humane gener koder for transkripsjons-regulatorer, men bare rundt 250 gjær-gener gjør det. Utviklingen av dyr domineres av celle-celle interaksjoner og av forskjellig genuttrykk.
iii. Den tredje klassen av ikke-kodende RNA har en mer usikker status: den inkluderer gener som koder for mikroRNA (miRNA); det er minst 500 av disse hos mennesker. Sammen med de regulatoriske proteinene spiller de en viktig rolle i å kontrollere genuttrykk under dyrs utvikling, men hele omfanget av deres betydning er fremdeles uklar.
Kromatin dans i cellekjernen gjennom ekstensile motorer
Transkripsjon og genregulering Genom-topologi har dukket opp som en nøkkelaktør i alle genomfunksjoner. Selv om et bidrag fra lokal genomsløyfing i transkripsjon lenge har blitt verdsatt, har nyere observasjoner avdekket viktigheten av langvarige interaksjoner, og genom-omfattende studier har avdekket den universelle karakteren av slike regulatoriske genom-topologiske interaksjoner i genregulering. Flere typer kromosomale interaksjoner, enten i form av løkker mellom sekvenser på de samme kromosomene eller inter-kromosomale interaksjoner, har vist seg som viktige mekanismer i genregulering.
Arten av genomtopologi er veldig presis, og så er dens regulatoriske funksjoner i genuttrykk og vedlikehold av genom. Det fremvoksende bildet er en av omfattende selvhånderende tilbakemeldinger mellom aktivitet og romlig organisering av genomet, noe som tyder på et selvorganiserende og selvutviklende system som bruker epigenetisk dynamikk for å regulere genomfunksjonen, som svar på regulatoriske signaler og for å forplante celle-skjebne hukommelse.
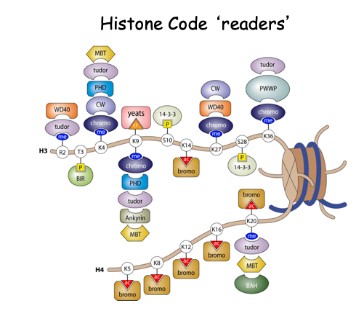
Post-transkripsjonelle modifikasjoner (PTM) av histoner påvirker gentranskripsjonen
I henhold til histonkode-hypotesen, blir mønsteret for histonmodifisering gjenkjent av proteiner, omtrent som et språk eller kode. Ett mønster av histon-modifisering kan tiltrekke proteiner som hemmer transkripsjon. Alternativt kan en annen kombinasjon av histon-modifikasjoner tiltrekke proteiner, slik som ATP-avhengige kromatin-ombyggings-komplekser, som fremmer gentranskripsjon. På denne måten spiller histonkoden en nøkkelrolle i tilgangen til informasjonen i genomene til eukaryote arter. Post-translasjonelle modifikasjoner (PTM) av histoner gir en finjustert mekanisme for å regulere kromatinstruktur og dynamikk. I tillegg til kombinatoriske PTM-er som fungerer både synergistisk og antagonistisk, er det nå en forståelse for PTM-asymmetri innen individuelle nukleosomer, nye typer PTM-er med unike funksjoner, nukleosomer som har histonvarianter, og kjerne-kompartmentaliserings hendelser som alle bidrar til det endelige resultatet av kromatinorganisasjon og funksjon
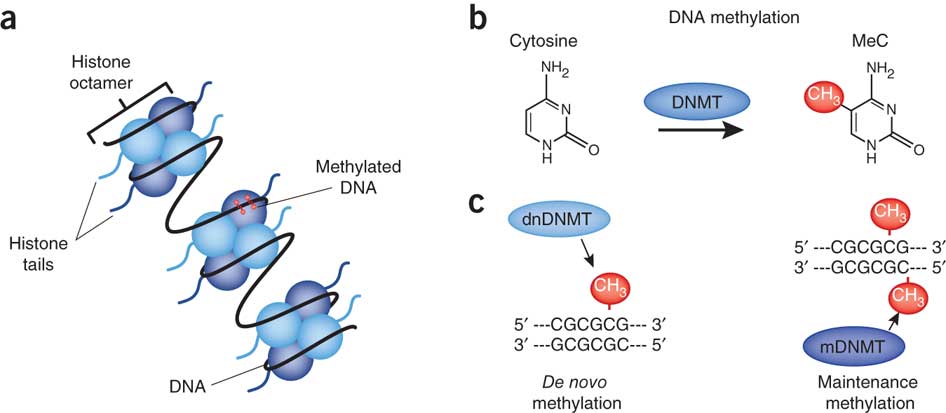 DNA-metyleringskoden og språket
DNA-metyleringskoden og språket
En av de mest kjente epigenetiske mekanismene er DNA-metylering der et lite molekyl (en metylgruppe) blir tilsatt til DNA-makromolekylet på bestemte steder. Som en strekkode eller markør indikerer metylgruppen for eksempel hvilke gener i DNA som skal slås på. Denne DNA-metyleringen oppnås via virkningen av en proteinmaskin som tilfører metylgruppen på nøyaktig riktig sted i DNA-strengen. Metylering av DNA skjer på visse målsteder langs DNA-sekvensen der spesifikke korte DNA-sekvenser vises. Disse sekvensene blir funnet av spesielle proteiner når de beveger seg langs DNA. De spesielle proteinene søker etter disse sekvensene og tilfører en metylgruppe til adeninbasen som vises i sekvensen. Proteinet binder seg til DNA, vrir heliksen slik at adeninbasen roterer i en nøyaktig formet lomme i proteinet, og proteinet letter deretter overføringen av metylgruppen fra et kort donormolekyl for eksempel til adenin.
Homeobox og Hox gener
Denne familien av beslektede gener bestemmer kroppens form. Det deler opp embryoet langs hode-til-sete-aksen i felt av celler som til slutt blir lemmer og andre strukturer. Starter som et befruktet egg med et homogent utseende, oppstår et embryo laget av hud, muskler, nerver og annet vev gradvis gjennom delingen av celler. Lenge før de fleste celler i det fremvoksende legeme begynner å spesialisere seg, etableres imidlertid en plan som utpeker hovedregioner i kroppen - hodet, mageregionen, setet og så videre. Denne planen hjelper tilsynelatende identiske kombinasjoner av vev med å ordne seg i tydelig forskjellige anatomiske strukturer, for eksempel armer og ben. Individuelle gener formidler noen av de utviklingsmessige beslutningene som er involvert i å etablere den embryonale kroppsplanen.
 "Junk DNA"
"Junk DNA"
MicroRNAs - En gang avskrevet som 'Junk' - Er bekreftet å ha viktige genetiske reguleringsfunksjon I 2008 bemerket Scientific American at microRNAs "en gang ble avskjediget som søppel" og sa følgende:
Små utdrag av genomet kjent som microRNA ble lenge antatt å være genomisk avfall fordi de ble transkribert fra såkalt "søppel-DNA", deler av genomet som ikke har informasjon for å lage proteiner ansvarlige for forskjellige cellefunksjoner. Det har blitt bygd bevis siden 1993, men at microRNA er alt annet enn genetisk avfall. Tvert imot, forskere sier at det faktisk spiller en avgjørende rolle i å slå protein-kodende gener på eller av og regulere mengden protein disse genene produserer.
Transposons og Retrotransposons
Slående bevis har samlet seg som indikerer at noen provirale sekvenser og HERV-proteiner til og med kan tjene vertenes behov, og derfor er under positiv seleksjon. Den bemerkelsesverdige fremgangen i analysen av vertsgenomer har brakt fram den betydelige effekten av HERVs og andre retroelementer på genetisk variasjon, genomutvikling og genregulering.
Centrosomer
Sentrosomer spiller en sentral rolle i utviklingen: Et froskeegg kan bli indusert til å utvikle seg til en frosk bare ved å injisere en sæd-centrosom - ingen sæd-DNA er nødvendig. En annen ikke-genetisk faktor som er involvert i utviklingen er eggcellens membranmønster.
Cytoskeletale matriser
Det nøyaktige arrangementet av mikrotubuli i cytoskjelettet utgjør en form for kritisk strukturell informasjon. Verken tubulin-underenhetene eller genene som produserer dem, står for forskjellene i formen til mikrotubulær-matriser som skiller forskjellige typer embryoer og utviklingsveier. I stedet er strukturen til selve mikrotubularsystemet, nok en gang, bestemt av plasseringen og arrangementet av dens underenheter, ikke egenskapene til underenhetene selv. Jonathan Wells forklarer det på denne måten: "Det som betyr noe i  [embryologisk] utvikling er formen og beliggenheten til mikrotubule-matriser, og formen og beliggenheten til en mikrotubule-gruppe er ikke bestemt av enhetene." Rettet transport involverer cytoskjelettet, men det avhenger også av romlig lokaliserte mål i membranen som er på plass før transport skjer. Utviklingsbiologer har vist at disse membranmønstrene spiller en avgjørende rolle i den embryologiske utviklingen av fruktfluer.
[embryologisk] utvikling er formen og beliggenheten til mikrotubule-matriser, og formen og beliggenheten til en mikrotubule-gruppe er ikke bestemt av enhetene." Rettet transport involverer cytoskjelettet, men det avhenger også av romlig lokaliserte mål i membranen som er på plass før transport skjer. Utviklingsbiologer har vist at disse membranmønstrene spiller en avgjørende rolle i den embryologiske utviklingen av fruktfluer.
Signalering mellom celler orienterer den mitotiske spindelen
Hos flercellede dyr tjener cellekommunikasjon noen ganger til å orientere retningen som celler deler seg i. Kontroll av delingsorientering er foreslått å være kritisk for å dele opp utviklingsdeterminanter og for å opprettholde epitelarkitektur. Overraskende nok er det få tilfeller der vi forstår mekanismene som eksterne signaler, overført ved intercellulær signalering, spesifiserer delingsorientering av dyreceller. Man kan forutsi at cytosoliske molekyler eller komplekser eksisterer som er i stand til å tolke ekstrinsiske signaler, og oversette posisjonene til disse ledningene til krefter på mikrotubuli i den mitotiske spindelen.
I løpet av de siste årene har det blitt identifisert et sentralt intracellulært kompleks som er nødvendig for å trekke krefter på mitotiske spindler i Drosophila, C. elegans og virveldyrsystemer. Et medlem av dette komplekset, et protein med TetratricoPeptid Repeterende (TPR) og GoLoco (Ga-bindende) domener, er funnet lokalisert i posisjoner som sammenfaller med stillingene til spindelorienterende ekstracellulære signaler. Fungerer TPR-GoLoco proteiner som konserverte, romlig regulerte formidlere av spindelorientering ved intercellulær signalering? Her gjennomgår vi relevant bevis blant tilfeller fra forskjellige dyre-systemer der dette proteinkomplekset har funnet å lokalisere til spesifikke celle-cellekontakter og å være involvert i å orientere mitotiske spindler.
Membranmål
Preekisterende membranmål, allerede plassert på innsiden av overflaten av eggcellen, bestemmer hvor disse molekylene vil feste seg og hvordan de vil fungere. Disse membranmålene gir viktig informasjon - romlige koordinater - for embryologisk utvikling.
Ionekanaler og elektromagnetiske felt
 Eksperimenter har vist at elektromagnetiske felt har 'morfogenetiske' effekter - med andre ord effekter som påvirker formen til en utviklende organisme. Spesielt har noen eksperimenter vist at den målrettede forstyrrelsen av disse elektriske feltene forstyrrer normal utvikling på måter som antyder at feltene kontrollerer morfogenesen. Kunstig påførte elektriske felt kan indusere og lede celle-migrasjon. Det er også bevis på at likestrøm kan påvirke genuttrykk, noe som betyr at internt genererte elektriske felt kan gi romlige koordinater som styrer embryogenese. Selv om ionekanalene som genererer feltene består av proteiner som kan kodes av DNA (akkurat som mikrotubuli består av underenheter kodet av DNA), er mønsteret i membranen ikke. I tillegg til informasjonen i DNA som koder for morfogenetiske proteiner, påvirker den romlige ordningen og fordelingen av disse ionekanalene dyrets utvikling.
Eksperimenter har vist at elektromagnetiske felt har 'morfogenetiske' effekter - med andre ord effekter som påvirker formen til en utviklende organisme. Spesielt har noen eksperimenter vist at den målrettede forstyrrelsen av disse elektriske feltene forstyrrer normal utvikling på måter som antyder at feltene kontrollerer morfogenesen. Kunstig påførte elektriske felt kan indusere og lede celle-migrasjon. Det er også bevis på at likestrøm kan påvirke genuttrykk, noe som betyr at internt genererte elektriske felt kan gi romlige koordinater som styrer embryogenese. Selv om ionekanalene som genererer feltene består av proteiner som kan kodes av DNA (akkurat som mikrotubuli består av underenheter kodet av DNA), er mønsteret i membranen ikke. I tillegg til informasjonen i DNA som koder for morfogenetiske proteiner, påvirker den romlige ordningen og fordelingen av disse ionekanalene dyrets utvikling.
Sukkerkoden
Disse sekvensspesifikke informasjonsrike strukturer påvirker arrangementet av forskjellige celletyper under embryologisk utvikling. Således refererer noen cellebiologer nå til arrangementene av sukkermolekyler som 'sukkerkoden' og sammenligner disse sekvensene med den digitalt kodede informasjonen som er lagret i DNA. Som biokjemiker Hans-Joachim Gabius bemerker, gir sukker et system med 'høy tetthetskoding' som er "viktig for at celler skal kunne kommunisere effektivt og raskt gjennom komplekse overflateinteraksjoner." I følge Gabius "overgår disse [sukker] -molekylene aminosyrer og nukleotider langt i informasjons-lagringskapasitet." Så de presist arrangerte sukkermolekylene på overflaten av celler representerer tydelig en annen informasjonskilde uavhengig av den som er lagret i DNA-basesekvenser. Disse kaskadene er sammen med selve cellebegivenheten assosiert med den 'kodende informasjonen' på en celleoverflate, eller ved bruk av en annen terminologi, realiseres på grunn av en instruksjon for cellen fra det morfogenetiske feltet til en organisme. De konkrete signaloverføringsveiene som forbinder 'kodingsinformasjonen' på en celleoverflate og uttrykket av de gitte settene med gener, må belyses.
 Over og utover: epigenetisk informasjon
Over og utover: epigenetisk informasjon
Gener alene bestemmer ikke den tredimensjonale formen og strukturen til et dyr. Spesielt utviklingsbiologer oppdager nå flere og flere måter hvor avgjørende informasjon for å bygge kroppsplaner, formidles av formen og strukturen til embryonale celler, inkludert informasjon fra både det ubefruktede og befruktede egget. DNA hjelper direkte proteinsyntese. Deler av DNA-molekylet er også med på å regulere tidspunkt og uttrykk for genetisk informasjon og syntese av forskjellige proteiner i celler. Likevel når proteiner er syntetisert, må de ordnes i høyere nivåer av proteiner og strukturer.
Den tredimensjonale strukturen eller den romlige arkitekturen til embryonale celler spiller viktige roller for å bestemme kroppsplandannelse under embryogenese. Utviklingsbiologer har identifisert flere kilder til epigenetisk informasjon i disse cellene.
Stephen C. Meyer, Darwins Doubt: Neo darwinisme og utfordringen med epigenetisk informasjon
Disse forskjellige kildene til epigenetisk informasjon i embryonale celler utgjør en enorm utfordring for tilstrekkeligheten av den neo-darwinistiske mekanismen. I følge neo-Darwinisme oppstår ny informasjon, form og struktur fra naturlig seleksjon som virker på tilfeldige mutasjoner, som oppstår på et veldig lavt nivå i det biologiske hierarkiet - innenfor den genetiske teksten. Likevel er både kroppsplandannelse under embryologisk utvikling og større morfologisk innovasjon i løpet av livshistorien avhengig av en spesifisitet av ordningen på et mye høyere nivå i organisasjonshierarkiet, et nivå som DNA alene ikke bestemmer.
Hvis DNA ikke er helt ansvarlig for hvordan et embryo utvikler seg - for morfogenese av kroppsplan - kan DNA-sekvenser muteres på ubestemt tid og fremdeles ikke produsere en ny kroppsplan, uavhengig av hvor mye tid og antall mutasjonsforsøk som er tilgjengelig for evolusjonsprosess. Genetiske mutasjoner er rett og slett feil verktøy for den aktuelle jobben. Selv i et beste scenario - et som ignorerer den enorme usannsynligheten ved å generere nye gener ved mutasjon og seleksjon - ville mutasjoner i DNA-sekvens bare produsere ny genetisk informasjon. Men å bygge en ny kroppsplan krever mer enn bare genetisk informasjon. Det krever både genetisk og epigenetisk informasjon - informasjon som pr. definisjon ikke lagres i DNA og dermed ikke kan genereres av mutasjoner til DNA. Det følger at mekanismen for naturlig seleksjon som virker på tilfeldige mutasjoner i DNA ikke i seg selv kan generere nye kroppsplaner, slik som de som først oppstod i den kambriske eksplosjonen.
 Prinsipiell betydning av evolusjon i lærebøker -
Hva er faktum:
Prinsipiell betydning av evolusjon i lærebøker -
Hva er faktum:
1. Endring over tid; naturens historie; enhver hendelsesrekkefølge i naturen
2. Endringer i frekvensene av alleler i genpoolen til en populasjon
3. Begrenset felles avstamning: ideen at bestemte grupper av organismer har stammet fra en vanlig stamfar.
4. Mekanismene som er ansvarlige for endringen som kreves for å produsere begrenset nedstigning med endring; hovedsakelig naturlig utvalg som handler på tilfeldige variasjoner eller mutasjoner
Hva er ikke faktum:
5. Universell felles avstamming: ideen om at alle organismer er nedstammet fra en felles stamfar.
6. Blind urmakeroppgave: ideen om at alle organismer har nedstammet fra felles forfedre gjennom ikke-ledede, uintelligente, formålsløse, materielle prosesser lik naturlige utvalg som handler på tilfeldige variasjoner eller mutasjoner; ideen om at den darwinistiske mekanismen for naturlig seleksjon som virker på tilfeldig variasjon, og andre tilsvarende naturalistiske mekanismer, er helt tilstrekkelig for å forklare opprinnelsen til nye biologiske former og designets utseende i komplekse organismer.
1. Neo-Darwinisme og den moderne syntese foreslår et gensentrisk syn, et vitenskapelig metabiologisk forslag som går tilbake til Darwins landemerke-bok 'Om artenes opprinnelse' i 1859, der først naturlig utvalg ble foreslått som mekanisme for biologisk mangfold, og senere, genvariasjon som definerer hvordan kropper bygges og organiseres.
2. Forskningsforskere har oppdaget at robuste nettverk av interaksjoner og biologisk funksjon, større morfologisk innovasjon, utvikling og kroppsform er basert på integrerende mekanismer, samspillet mellom gener med det genregulerende nettverket, Transposons og Retrotransposons, såkalt Junk DNA, spleising, og over et dusin epigenetiske koder, membranmål og mønstre, cytoskeletale matriser, sentrosomer, ionekanaler, sukkermolekyler på utsiden av celler (sukkerkoden), som ikke er spesifisert av kjernefysisk DNA - det vil si arv er ikke definert gjennom DNA-sekvenser alene.
3. Vitenskapen kommer til å erkjenne at ingen av de nylig foreslåtte alternativene, som den tredje veien, Saltationisme, Saltatorisk ontogeni, mutationisme, Genetisk drift, eller kombinerte teorier, utøver full rettferdighet ved å ta hensyn til alt organisatorisk biofysiologisk hierarki og kompleksitet som empirisk vitenskapen har kommet til å oppdage. Som sådan er det kun et helhetlig syn, strukturalisme, som tar hensyn til alle påvirkninger som danner celleform og størrelse, kroppsutvikling og vekst, og utøver rettferdighet for det vitenskapelige beviset.
4. å undersøke hvilke som til slutt svarer for den kompleksiteten som oppdages i livet, er bare tilfredsstillende, når det epistemologisk mangelfulle grunnlaget for metodologisk naturalisme er tatt ut av esken, og erstattet av et nytt paradigme, der alle mulige mekanismer og årsaksmessige påvirkninger tillates å være gransket, undersøkt og vitenskapelig testet, inkludert samspillet til og den kreative kraften ved en ekstern intelligent, mental agent, utenfor den kjente fysiske verden, som gjennom sin transcendente kraft skaper, former og bygger alle fysiobiologiske livsformer i all sin forbløffende mangfold.
 Hvis mekanismene som foreslått av den moderne utvidede evolusjonære syntesen er utilstrekkelige, må det finnes ut hvilke ANDRE mekanismer som definerer kroppsarkitektur, vekst, differensiering, fenotype og kroppsform, og DET kan evalueres, hvordan opphavet til disse mekanismer blir best forklart. Når det kommer ut, er en STOR bidragsyter til å forklare livet og dets kompleksitet INFORMASJON. Instruksjonsmessig, kompleks, spesifisert, kodifisert informasjon som fungerer som en blåkopi (gener), men også informasjon som er PRE-PROGRAMMERET og lagret i epigenetiske cellefunksjoner på strukturelt nivå, som er forhåndsinstruert til å svare på miljømessige signaler, utvikling og ernæringskrav, og de er egnet til å kommunisere, krysskoble, signalisere, regulere, styre, kontrollere, rekruttere, tolke, gjenkjenne, orkestrere, utdype strategier, veilede og så videre.
Hvis mekanismene som foreslått av den moderne utvidede evolusjonære syntesen er utilstrekkelige, må det finnes ut hvilke ANDRE mekanismer som definerer kroppsarkitektur, vekst, differensiering, fenotype og kroppsform, og DET kan evalueres, hvordan opphavet til disse mekanismer blir best forklart. Når det kommer ut, er en STOR bidragsyter til å forklare livet og dets kompleksitet INFORMASJON. Instruksjonsmessig, kompleks, spesifisert, kodifisert informasjon som fungerer som en blåkopi (gener), men også informasjon som er PRE-PROGRAMMERET og lagret i epigenetiske cellefunksjoner på strukturelt nivå, som er forhåndsinstruert til å svare på miljømessige signaler, utvikling og ernæringskrav, og de er egnet til å kommunisere, krysskoble, signalisere, regulere, styre, kontrollere, rekruttere, tolke, gjenkjenne, orkestrere, utdype strategier, veilede og så videre.
1. Celler bruker sofistikerte informasjons-overførings- og forsterknings-systemer (signalveier), informasjonstolkning, kombinasjon og seleksjon (Gene-regulatorisk nettverk) koding og transkripsjon (DNA og RNA-polymerasemaskiner) transmisjons- (mRNA) og dekodings-systemer (ribosomer).
2. Oppsett av informasjonsoverføringssystemer, ala. overføring, forsterkning, tolkning, kombinasjon, valg, koding, overføring og dekoding er alltid en bevisst handling
3. Eksistensen av det genetiske informasjons-overføringssystemet forklares best ved implementeringen av en intelligent designer.
Oversettelse og bilder ved Asbjørn E. Lund